Механизм действия гормонов гипофиза и гипоталамуса
Гормональная регуляция начинается с процесса синтеза и секреции гормонов в железах внутренней секреции. Они функционально взаимосвязаны и представляют единое целое. Процесс биосинтеза гормонов, осуществляемый в специализированных клетках, протекает спонтанно и закреплен генетически. Генетический контроль биосинтеза большинства белково-пептидных гормонов, в частности аденогипофизотропных, осуществляется чаще всего непосредственно в полисомах гормонов-предшественников или на уровне образования мРНК самого гормона, тогда как биосинтез гормонов гипоталамуса осуществляется путем образования мРНК белков-ферментов, регулирующих различные этапы образования гормона, т. е. происходит внерибосомальный синтез. Формирование первичной структуры гормонов белково-пептидной природы - результат прямой трансляции нуклеотидных последовательностей соответствующих мРНК, синтезируемых на активных участках генома гормонпродуцирующих клеток. Структура большинства белковых гормонов или их предшественников формируется в полисомах по общей схеме биосинтеза белка. Отметим, что возможность синтеза и трансляции мРНК данного гормона или его предшественников специфична для ядерного аппарата и полисом определенного типа клеток. Так, СТГ синтезируется в малых эозинофилах аденогипофиза, пролактин - в больших эозинофильных, а гонадотропины - в особых базофильных клетках. Несколько иначе происходит биосинтез ТРГ и ЛГ-РГ в клетках гипоталамуса. Эти пептиды образуются не в полисомах на матрице мРНК, а в растворимой части цитоплазмы под влиянием соответствующих синтетазных систем.
Прямая трансляция генетического материала в случаях выделения большинства полипептидных гормонов часто приводит к образованию малоактивных предшественников - полипептидных препрогормонов (прегормонов). Биосинтез полипептидного гормона складывается из двух различных этапов: рибосомального синтеза неактивного предшественника на матрице мРНК и посттрансляционного образования активного гормона. Первый этап протекает обязательно в клетках аденогипофиза, второй же может осуществляться и вне его.
Посттрансляционная активация гормональных предшественников возможна двумя способами: путем многоступенчатой ферментативной деградации молекул транслируемых крупномолекулярных предшественников с уменьшением размеров молекулы активируемого гормона и за счет неферментативной ассоциации про-гормональных субъединиц с укрупнением размеров молекулы активируемого гормона.
В первом случае посттрансляционная активация характерна для АКТУ, бета-липотропина, а во втором - для гликопротеидных гормонов, в частности гонадотропинов и ТТГ.
Последовательная активация белково-пептидных гормонов имеет прямой биологический смысл. Во-первых, при этом ограничиваются гормональные эффекты в месте образования; во-вторых, обеспечиваются оптимальные условия для проявления полифункциональных регуляторных эффектов при минимальном использовании генетического и строительного материала, а также облегчается клеточный транспорт гормонов.
Выделение гормонов происходит, как правило, спонтанно, причем не непрерывно и равномерно, а импульсивно, отдельными дискретными порциями. Это обусловлено, по-видимому, циклическим характером процессов биосинтеза, внутриклеточного депонирования и транспорта гормонов. В условиях физиологической нормы секреторный процесс должен обеспечить определенный базальныи уровень гормонов в циркулирующих жидкостях. Этот- процесс, как и биосинтез, находится под контролем специфических факторов. Секреция гипофизарных гормонов в первую очередь определяется соответствующими рилизинг-гормонами гипоталамуса и уровнем циркулирующих гормонов в крови. Образование же самих гипоталамических рилизинг-гормонов зависит от влияния нейромедиаторов адренергической или холинергической природы, а также концентрации гормонов желез-«мишеней» в крови.
Биосинтез и секреция тесно взаимосвязаны. Химическая природа гормона и особенности механизмов его секреции обусловливают степень сопряженности этих процессов. Так, этот показатель максимален в случае секреции стероидных гормонов, которые относительно свободно диффундируют через клеточные мембраны. Величина сопряженности биосинтеза и секреции белково-пептидных гормонов и катехоламинов минимальна. Эти гормоны освобождаются из клеточных секреторных гранул. Промежуточное положение по этому показателю занимают тиреоидные гормоны, которые секретируются путем освобождения их из белково-связанной формы.
Таким образом, следует подчеркнуть, что синтез и секреция гормонов гипофиза и гипоталамуса осуществляются в определенной степени раздельно.
Главным структурно-функциональным элементом секреторного процесса белково-пептидных гормонов являются секреторные гранулы или везикулы. Это особые морфологические образования овоидной формы различного размера (100-600 нм), окруженные тонкой липопротеидной мембраной. Секреторные гранулы гормонпродуцирующих клеток возникают из комплекса Гольджи. Его элементы окружают прогормон или гормон, постепенно формируя гранулы, которые выполняют ряд взаимосвязанных функций в системе процессов, обусловливающих секрецию гормонов. Они могут быть местом активации пептидных прогормонов. Вторая функция, которую выполняют гранулы, - хранение гормонов в клетке до момента воздействия специфического секреторного стимула. Мембрана гранул ограничивает выход гормонов в цитоплазму и защищает гормоны от действия цитоплазматических ферментов, способных их инактивировать. Определенное значение в механизмах депонирования имеют особые вещества и ионы, содержащиеся внутри гранул. К ним относятся белки, нуклеотиды, ионы, главное назначение которых - образование нековалентных комплексов с гормонами и предотвращение проникновения их через мембрану. Секреторные гранулы обладают еще одним очень важным качеством - способностью перемещаться к периферии клетки и транспортировать депонированные в них гормоны к плазматическим мембранам. Движение гранул осуществляется внутри клеток при участии органелл клетки - микрофиламентов (их диаметр 5 нм), построенных из белка актина, и полых микротрубок (диаметр 25 нм), состоящих из комплекса сократительных белков тубулина и динеина. В случае необходимости блокады секреторных процессов обычно применяют препараты, разрушающие микрофиламенты или диссоциацию микротрубок (цитохалазин В, колхицин, винбластин). Внутриклеточный транспорт гранул требует затрат энергии и присутствия ионов кальция. Мембраны гранул и плазматические мембраны при участии кальция вступают между собой в контакт, и секрет выделяется во внеклеточное пространство через «поры», образующиеся в клеточной мембране. Этот процесс называется экзоцитозом. Опустошенные гранулы способны в некоторых случаях реконструироваться и возвращаться в цитоплазму.
Пусковым моментом в процессе секреции белково-пептидных гормонов является повышенное образование АМФ (цАМФ) и увеличение внутриклеточной концентрации ионов кальция, которые проникают через плазматическую мембрану и стимулируют переход гормональных гранул к клеточной мембране. Описанные выше процессы регулируются как внутриклеточно, так и внеклеточно. Если внутриклеточная регуляция и саморегуляция гормонпродуцирующеи функции клеток гипофиза и гипоталамуса в значительной степени ограничены, то системные механизмы контроля обеспечивают функциональную активность гипофиза и гипоталамуса в соответствии с физиологическим состоянием организма. Нарушение регуляторных процессов может привести к серьезной патологии функций желез и, следовательно, всего организма.
Регуляторные влияния можно подразделить на стимулирующие и тормозящие. В основе всех регуляторных процессов лежит принцип обратных связей. Ведущее место в упорядочивании гормональных функций гипофиза принадлежит структурам ЦНС, и в первую очередь гипоталамусу. Таким образом, физиологические механизмы контроля деятельности гипофиза можно подразделить на нервные и гормональные.
Рассматривая процессы регуляции синтеза и секреции гипофизарных гормонов, следует прежде всего указать на гипоталамус с его способностью синтезировать и секретировать нейрогормоны - рилизинг-гормоны. Как указывалось, регуляция аденогипофизарных гормонов осуществляется с помощью рилизинг-гормонов, синтезируемых в определенных ядрах гипоталамуса. Мелкоклеточные элементы этих структур гипоталамуса имеют проводящие пути, контактирующие с сосудами первичной капиллярной сети, через которые и поступают рилизинг-гормоны, достигая аденогипофизарных клеток.
Рассматривая гипоталамус как нейроэндокринный центр, т. е. как место трансформации нервного импульса в специфический гормональный сигнал, носителем которого являются рилизинг-гормоны, ученые исследуют возможность влияния различных медиаторных систем непосредственно на процессы синтеза и секреции аденогипофизарных гормонов. С помощью усовершенствованных методических приемов исследователи выявили, например, роль дофамина в регуляции секреции ряда тропных гормонов аденогипофиза. В данном случае дофамин выступает не только как нейромедиатор, упорядочивающий функцию гипоталамуса, но и в качестве рилизинг-гормона, принимающего участие в регуляции функции аденогипофиза. Аналогичные данные получены и в отношении норадреналина, участвующего в.контроле секреции АКТГ. Факт двойного контроля синтеза и секреции адено-гипофизотропных гормонов в настоящее время установлен. Основной же точкой приложения различных нейромедиаторов в системе регуляции гипоталамических рилизинг-гормонов являются структуры гипоталамуса, в которых они синтезируются. В настоящее время спектр физиологически активных веществ, причастных к регуляции гипоталамических нейрогормонов, достаточно широк. Это классические нейромедиаторы адренергической и холинергической природы, ряда аминокислот, вещества с морфиноподобным действием - эндорфины и энкефалины. Эти вещества являются основным связующим звеном между ЦНС и эндокринной системой, что в конечном итоге обеспечивает их единство в организме. Функциональная активность гипоталамических нейроэндокринных клеток может непосредственно контролироваться различными отделами головного мозга с помощью нервных импульсов, поступающих по различным афферентным путям.
В последнее время в нейроэндокринологии возникла еще одна проблема - изучение функциональной роли рилизинг-гормонов, локализованных в других структурах ЦНС, за пределами гипоталамуса и не имеющих прямого отношения к гормональной регуляции аденогипофизарных функций. Экспериментально подтверждено, что они могут рассматриваться и как нейромедиаторы, и как нейромодуляторы ряда системных процессов.
В гипоталамусе рилизинг-гормоны локализованы в определенных областях или ядрах. Так, например, ЛГ-РГ локализован в переднем и медиобазальном гипоталамусе, ТРГ - в области среднего гипоталамуса, КРГ - в основном в его задних отделах. Это не исключает также и диффузного распределения в железе нейрогормонов.
Основная функция аденогипофизарных гормонов сводится к активации ряда периферических эндокринных желез (коры надпочечников, щитовидной железы, гонад). Тропные гормоны гипофиза - АКТГ, ТТГ, ЛГ и ФСГ, СТГ - вызывают специфические ответные реакции. Так, первый вызывает разрастание (гипертрофию и гиперплазию) пучковой зоны адреналовой коры и усиление в ее клетках синтеза глюкокортикоидов; второй - главный регулятор морфогенеза фолликулярного аппарата щитовидной железы, различных стадий синтеза и секреции тиреоидных гормонов; ЛГ - основной стимулятор овуляции и образования желтого тела в яичниках, роста интерстициальных клеток в семенниках, синтеза эстрогенов, прогестинов и гонадальных андрогенов; ФСГ вызывает ускорение роста овариальных фолликулов, сенсибилизирует их к действию ЛГ, а также активирует сперматогенез; СТГ, действуя стимулирующим образом на выделение печенью соматомединов, определяет линейный рост организма и анаболических процессов; ЛТГ способствует проявлению действия гонадотропинов.
Следует также отметить, что тропные гормоны гипофиза, проявляя свое действие как регуляторы функций периферических эндокринных желез, часто способны оказать прямой эффект. Так, например, АКТГ как главный регулятор синтеза глюкокортикоидов дает ряд экстраадреналовых эффектов, в частности липолитический и меланоцитостимулирующий.
Гормоны гипоталамо-гипофизарного происхождения, т. е. белково-пептидные, очень быстро исчезают из крови. Период их полураспада не превышает 20 мин и в большинстве случаев продолжается 1-3 мин. Белково-пептидные гормоны быстро накапливаются в печени, где происходит их интенсивная деградация и инактивация под действием специфических пептидаз. Этот процесс может наблюдаться и в других тканях, а также в крови. Метаболиты белково-пептидных гормонов выводятся, по-видимому, преимущественно в форме свободных аминокислот, их солей и небольших пептидов. Они экскретируются в первую очередь с мочой и желчью.
Гормоны чаще всего обладают достаточно выраженной тропностью физиологического действия. Например, АКТГ действует на клетки коры надпочечников, жировой ткани, нервной ткани; гонадотропины - на клетки гонад, гипоталамуса и ряд других структур, т. е. на органы-, ткани-, клетки-«мишени». Гормоны гипофиза и гипоталамуса обладают широким спектром физиологического действия на клетки разного типа и на различные обменные реакции в одних и тех же клетках. Структуры организма по степени зависимости их функций от действия тех или иных гормонов подразделяются на гормонзависимые и гормончувствительные. Если первые полностью обусловлены наличием гормонов в процессе полноценной дифференцировки и функционирования, то гормончувствительные клетки отчетливо проявляют свои фенотипические признаки и без соответствующего гормона, степень проявления которых модулируется им в разном диапазоне и определяется за счет наличия особых рецепторов у клетки.
Взаимодействие гормонов с соответствующими рецепторными белками сводится к нековалентному, обратимому связыванию гормональных и рецепторных молекул, в результате чего образуются специфические белок-лигандные комплексы, способные включать множественные гормональные эффекты в клетке. Если рецепторный белок в ней отсутствует, то она резистентна к действию физиологических концентраций гормона. Рецепторы являются необходимыми периферическими представителями соответствующей эндокринной функции, обусловливающими исходную физиологическую чувствительность реагирующей клетки к гормону, т. е. возможность и интенсивность приема, проведения и реализации гормонального синтеза в клетке.
Эффективность гормональной регуляции клеточного метаболизма определяется как количеством активного гормона, поступающего в клетку-«мишень», так и уровнем содержания рецепторов в ней.
Медицинский эксперт-редактор
Портнов Алексей Александрович
Образование: Киевский Национальный Медицинский Университет им. А.А. Богомольца, специальность - "Лечебное дело"
Другие статьи по теме
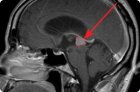
Гормонально-активная опухоль передней доли гипофиза – это пролактинома. Рассмотрим причины ее появления, факторы риска, симптомы, методы диагностики и лечения.
